
Introduction
Striped catfish (Pangasianodon hypophthalmus) is an important fish species providing a significant portion of human consumption. Similar to other economically important fish species, the production of striped catfish is becoming increasingly intensive, which poses numerous challenges, including the rapid increase in bacterial diseases, especially under suboptimal conditions such as high stocking density, poor water quality, and inadequate feeding practices (Hoa et al., 2021; Luu Tang Phuc et al., 2022; Nhu et al., 2019). Addressing these challenges, farmers frequently resort to therapeutic interventions involving the use of chemicals and antibiotics. However, these interventions have adverse effects on the environment and lead to concerns about residue accumulation in fish flesh. Additionally, the use of such chemicals may compromise both innate and adaptive immunity in fish, thereby posing potential risks to consumer health in the long term (Pepi & Focardi, 2021; Yuan et al., 2023). Consequently, the supplementation of bioactive compounds in diets to enhance the fish immune response is increasingly encouraged in aquaculture. Besides, biological compounds such as prebiotics have been reported to have positive effects on aquatic animals, such as enhancing growth, digestibility, and disease resistance (Amrullah & Wahidah, 2019; El-Nobi et al., 2021; Lu et al., 2019).
Prebiotics, including inulin, β-glucan, mannanoligosaccharide (MOS), arabinoxylan oligosaccharide (AXOS), oligosaccharides, galactooligosaccharide (GOS), and galactoglucomannans (GGM) serve as nourishment for beneficial bacteria within a host’s digestive tube (Wee et al., 2024). Additionally, prebiotics are characterized as substances capable of withstanding the acidic conditions in the stomach, undergoing fermentation by intestine microbiota, and fostering the growth of intestine microbiota to enhance host body health (Mugwanya et al., 2022; Wee et al., 2024). In essence, prebiotics are complex carbohydrates with long chains that serve as a source of energy for beneficial microorganisms or probiotics, thereby enhancing the overall health of an organism. Furthermore, Prebiotics contribute to inhibiting the adherence of pathogenic microorganisms to epithelial cells, facilitating the removal of detrimental bacteria from intestinal epithelial cells by acting as a receptor, modulating the host immune system, and controlling inflammation (Davani-Davari et al., 2019). Among them, peptidoglycan, one of the major components of the gram-positive bacteria cell wall (McDonald et al., 2005) is also reported to promote the feed utilization, host intestinal health, growth, and stimulate the immune system in human, terrestrial animals, shrimp, and fish species (Casadei et al., 2015; Pan et al., 2015; Zhou et al., 2006). Previous studies carried out on Japanese flounder (Zhou et al., 2006) and rainbow trout (Casadei et al., 2015) also reported that the beneficial effects of dietary peptidoglycan on fish growth, disease resistance, and immune responses at dose of more than 2 g/kg diet and 10 mg/kg diet, respectively. However, no results were recorded in striped catfish. The study was conducted with the hypothesis that peptidoglycan, given its positive impact on humans and terrestrial animals, would confer beneficial effects on fish. Therefore, the current study was conducted to assess the impacts of dietary supplementation of peptidoglycan extracted from Lactobacillus sp. on growth, feed utilization, and immune status in striped catfish.
Materials and Methods
The protocol of the experiment was accepted by the Animal Ethics Committee of Vietnam National University of Agriculture (Project No. I2-A-6670-1).
Striped catfish juveniles at size ~20 g from a local nursery farm were acclimatized in the closed tank system for two weeks. The environmental conditions were maintained similarly to the experiment. Fish were fed with commercial pellets for striped catfish (Aquaxcel, Cargill, 7414). After acclimatization, free disease and healthy fish were chosen for feeding trial.
Peptidoglycan compounds (Bio-Floc, Hanoi, Vietnam) were enzymatically hydrolyzed from Lactobacillus sp. (1012 CFU/g) The product was then lyophilized in powder form and stored in the freezer. The product was then suspended with a same volume of water (10 mL per 100 g of feed) at different levels of concentration: 0, 2.5, 5, and 10 g/kg of feed (named as PL0, PL2.5, PL5, and PL10 treatment). The experimental diets were daily prepared.
Healthy striped catfish juveniles (~26 g/fish) were randomly allocated into composite tanks of 250 L at a density of 25 fish per tank, 3 tanks per treatment. Fish were fed on the peptidoglycan-supplemented diets twice a day at 4% body weight. Fish were weighed after 2 weeks of experiment to adjust the feed amount. The experiment was conducted for 4 weeks. Tank system was continuously aerated and the environmental parameters such as temperature (ranging from 26°C to 29°C), pH (7.5 to 8), oxygen (6 to 8.5 mg/L), NH3/NH4+ (< 0.1 mg/L), and NO2 (< 0.1 mg/L) were monitored and maintained suitable to the requirements of striped catfish in which oxygen, pH, and temperature were measured two times per day and one time per week for other parameters. The feaces were daily siphoned.
On 14th and 28th day (T2 and T4) of the feeding trial, the fish blood samples were collected (3 fish per tank). Specifically, 1 mL of blood for each fish was collected and put in a heparin tube (30 µL of heparin in each tube) in which, 0.1 mL of fresh blood was be used for hematological analyses while 0.9 mL of heparin blood was centrifuged to collect the plasma for immune assays. The fish after sampling were removed from the experiment.
The hematological parameters, including hematocrit (HCT), total white blood cell count (WBC), monocytes, neutrophils, lymphocytes, and red blood cell count (RBC) were analyzed following the manufacturer’s protocols utilizing veterinary analyzers (URIT-3000 VETPLUS).
In a 96-well microplate, the lysozyme assay was started by mixing 10 μL of plasma with 130 μL of lyophilized Micrococcus lysodeikticus (0.6 mg/mL buffer, Sigma-Aldrich, MO, USA) suspension in phosphate buffer (pH 6.2). The absorbances at 450 nm measured every 5 minutes during an interval of 30 minutes were used to calculate units of lysozyme activity. The lysozyme activity unit (U/mL) is quantified as the enzyme amount inducing a reduction in absorbance of 0.001/min (Nhu et al., 2019).
Plasma sample was added into a 96-well plate, each sample was replicate in three. Water wells were used as the blank. Then, HBSS 1× (Thermo Fisher Scientific, USA) solution was added to a total volume of 75 μL for each well. A volume of 25 μL of reaction solution (TMB, Thermo Fisher Scientific, USA) was supplemented to each well. The mixture was then incubated for 2 min exactly. Finally, 25 µL of 2M H2SO4 (Sigma, USA) was added after incubation. The value of OD for each well was measured immediately by a spectrophotometer (Varioskan Lux, ThermoScientific) at 450 nm. Peroxidase activity is determined by the multiplication of the difference between the OD of each sample and blank with Df (Df = 1,000/sample volume used) and represented by U/mL (Nguyen et al., 2020).
After 4 weeks of experiment, the fish body weight, the number of fish were served to determine the fish growth, protein efficiency ratio (PER), feed conversion ratio (FCR), and survival rate. Moreover, the intestinal indices such as visceral somatic index (VSI), gastro-somatic index (GaSI), hepatosomatic index (HIS), and relative gut-length were calculated based on the weight and length of fish body, liver, and gut. The formula to calculate these parameters is described as follows:
Specific growth rate (SGR, %/day) = {[Ln (Final body weight) – Ln (Initial body weight)] / days of experiment} × 100
Daily weight gain (DWG, g/fish/day) = (Final body weight – Initial body weight) / days of experiment
Weight gain (WG, %) = ((FBW – IBW) / IBW) × 100
Feed conversion ratio (FCR) = Weight of feed intake / weight gain
Protein efficiency ratio (PER) = Weight gain / protein intake
Survival rate (%) = 100 × (final fish number / initial fish number)
Visceral somatic index (VSI, %) = (visceral weight / fish body weight) × 100
Gastro-somatic index (GaSI, %) = (gut-weight / fish body weight) × 100
Hepatosomatic index (HSI, %) = (liver weight / fish body weight) × 100
Relative gut-length (%) = (gut length / fish body length) × 100
Data will be subjected to one way ANOVA analyses with STATISTICA 10.0 software (Statsorf, Tulsa, OK, USA), followed by an LSD test using the diet replicate (n=3). Data was presented as means ± SD. Differences between groups are considered significant at p-value ≤ 0.05.
Results
At the beginning of experiment, the average body weight was homogenous approximately 26 g/fish. After 4 weeks of experiment, the final body weight ranged from 32.1 to 36.6 g/fish (Table 1). No significant differences between experiment treatments (p > 0.05) for growth variables such as DWG, SGR, and WG demonstrated that the supplementation of peptidoglycan did not influence the growth performance in striped catfish juveniles. Similarly, the survival rates were high (> 90%) and did not differ among the experiment groups (p > 0.05) indicating the rearing conditions were suitable for striped catfish and the enriched diet did not modify the survival rate of experimental fish. On the contrary, the significant impact (p < 0.05) of peptidoglycan supplementation was observed in variables of feed utilization (Table 1). Accordingly, the best values of PER and FCR were recorded in PL2.5-fed ones compared to other experimental groups, corresponding to 1.4 and 2.0, respectively.
PL0, PL2.5, PL5, PL10: commercial feed supplemented with peptidoglycan at 0, 2.5, 5, and 10 g/kg. IBW: initial body weight, FBW: final body weight, DWG: daily weight gain, SGR: specific growth rate, WG: weight gain. The data was represented by mean ± SD. Values with the different letters denote the significant difference (p < 0.05).
The intestinal parameters including VSI (ranged from 7.0% to 7.6%), HSI (from 2.4% to 2.6%), GaSI (from 2.6% to 3.0%), and relative gut length (from 106% to 143.8%) were recorded after 4 weeks of experiment and the results were presented in Table 2. Accordingly, the experimental treatments did not differ from all of the intestinal indices (p > 0.05), suggesting the supplementation of peptidoglycan in diet did not influence these studied parameters.
PL0, PL2.5, PL5, PL10: commercial feed supplemented with peptidoglycan at 0, 2.5, 5, and 10 g/kg. VSI: visceral somatic index, HSI: Hepatic somatic index, GaSI: Gastro-somatic index. The data was represented by mean ± SD. Values with the different letters denote the significant difference (p < 0.05).
The hematological indices such as WBC (ranging from 79.8 to 87.1 × 109 cells/L), RBC (from 2.0 to 2.4 × 1012 cells/L), lymphocytes (LYM, 30.2 to 35.5 × 109 cells/L), monocytes (MID, 26.9 to 30.3 × 109 cells/L), granulocytes (GRAN, 22.7 to 28.5 × 109 cells/L), and hematocrit (HCT, 25.3% to 30.7%) after 2 or 4 weeks did not differ between experiment treatments (Table 3). These results demonstrate that during 4 weeks of the experiment, the hematological parameters in striped catfish juveniles were not modified by the concentration of peptidoglycan supplementation in the diet as well as utilization duration.
PL0, PL2.5, PL5, PL10: commercial feed supplemented with peptidoglycan at 0, 2.5, 5, and 10 g/kg. WBC: White blood cells, RBC: red blood cells, LYM: lymphocytes, MID: monocytes, GRAN: granulocytes, HCT: hematocrit. T2 and T4 were samplings after 2 and 4 weeks of feeding trial. Data was presented as mean ± SD, the different letters denote the significant difference between treatments (p < 0.05).
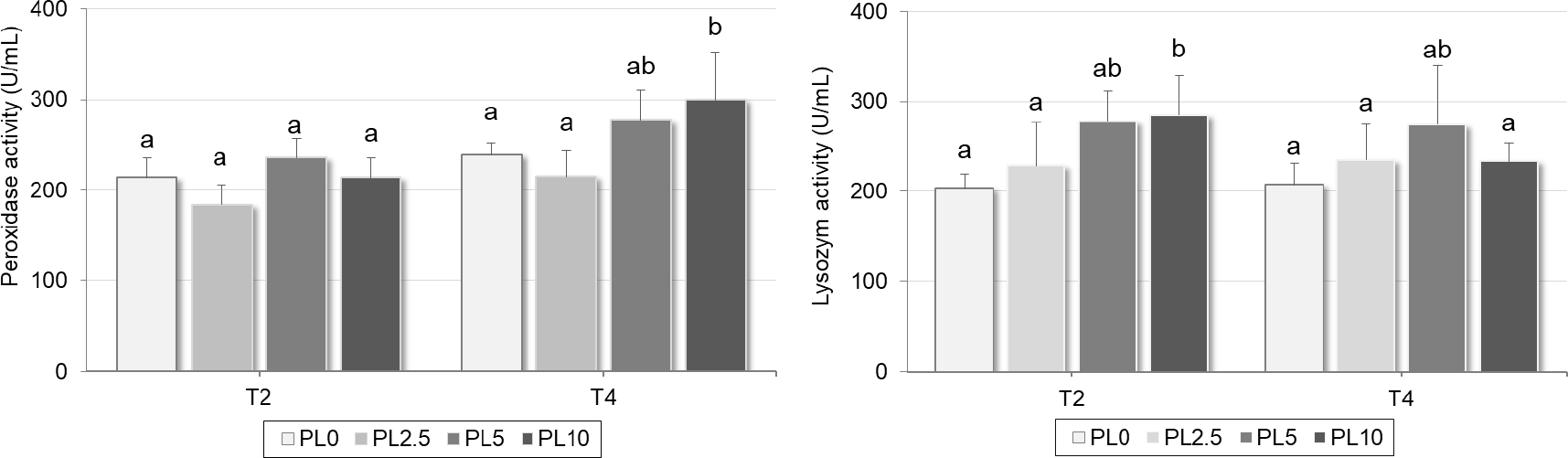
The immune parameters including peroxidase and lysozyme activities were also measured in fish blood plasma after 2 and 4 weeks, and the results were presented in Fig. 1. Accordingly, the peroxidase activity in blood plasma after 2 weeks of feeding trial ranged from 184.0 ± 28.7 to 235.6 ± 33.3 U/mL but no significant differences were found between treatments. The influences of dietary peptidoglycan were displayed after 4 weeks of feeding trial. Specifically, the highest value of peroxidase activity (p < 0.05) was recorded in PL10 treatment (300.0 ± 52.1 U/mL); higher than PL0 (238.6 ± 42.4 U/mL) and PL2.5 (215.1 ± 60.6 U/mL). Regarding on the results of lysozyme activity (Fig. 1), the influence of peptidoglycan supplementation in diet was observed at T2 instead of T4 like for peroxidase activity. However, a similar tendance was recorded for this immune variable when the highest value belonged to the PL10, and also higher than PL0 (207.5 ± 24.1 U/mL) and PL2.5 (235.4 ± 39.1 U/mL).
Discussion
Peptidoglycan is known as a prebiotics compound (Davani-Davari et al., 2019) that are metabolized by bacteria. The administration of prebiotic supplements often resulted in advantageous effects on fish growth or feed efficiency (Pan et al., 2015). The results with no favorable actions on these parameters were also recorded in fish (Yuji-Sado et al., 2015). Recent results published in Nile tilapia showed that diets enriched with peptidoglycan have improved fish growth, associated with an elevation in red blood cell count in the fish blood (Mai et al., 2023). The elevated count of red blood cells correspondingly led to increased oxygen levels in the fish bloodstream, consequently promoting higher growth performance in the fish (Ali & Mishra, 2022). The current study did not observe enhancements in fish growth in those fed with peptidoglycan-supplemented diets, nor did it observe an increase in red blood cells. This suggests a potential correlation between fish growth and this hematological parameter.
On the other hand, the best values of feed utilization including FCR and PER observed in PL2.5 could be explained by the feed digestion support effect of peptidoglycan in this study as well as previous descriptions of the effects of prebiotic products. Such beneficial outcomes could be attributed to alterations in the bacterial composition within the digestive tract, where prebiotics promote the activities of beneficial bacteria (Wee et al., 2024). The addition of peptidoglycan to the diet as a prebiotic generally regulates the native flora in the gastrointestinal tract by affecting enzyme activity. Dietary prebiotics can stimulate the secretion of digestive enzymes in fish intestines. Initially, dietary prebiotics contribute to the modulation of beneficial intestinal microflora, which is crucial for the secretion of digestive enzymes, particularly amylase (Munir et al., 2016). However, the treatments with higher doses did not induce the better results compared to low doses or control (Zhang et al., 2013). Thus, in the current experiment, the supplementation of peptidoglycan at 5 and 10 g/kg did not induce a favorable result compared to PL0. These obtained results may be explained by the interaction between beneficial bacteria and prebiotics in the host intestine. Prebiotics are polysaccharides that resist digestion by the host but are metabolized by bacteria. However, excessive levels of prebiotics in diets may not only foster gut microbiota but also potentially impede feed digestion (Baumgärtner et al., 2022).
Hematological parameters in fish offer insights into various aspects such as oxygen transport capacity, immune function, disease status, and nutritional status (Witeska et al., 2022). The principal indicators include red blood cells, white blood cells, and hematocrit. Red blood cells primarily function to transport oxygen to tissues (Stosik et al., 2020). The kidneys secrete a hormone known as erythropoietin, which stimulates the production of red blood cells. The concentration of this hormone rises in response to a deficiency in oxygen (Chen et al., 2017). In the current study, the total number of red blood cells in fish fed with peptidoglycan-supplemented diets was comparable to that in the control group without peptidoglycan supplementation. This result may be attributed to the efficient digestion of peptidoglycan by the bacterial population in the intestine of striped catfish. Consequently, the residual compounds, such as exogenous compounds, were insufficient to stimulate blood circulation and increase red blood cell counts. Thus, the results indicate that peptidoglycan supplementation at 2.5 g/kg of feed led to improved feed utilization but did not enhance the growth of striped catfish juveniles.
Conversely, white blood cells play a critical role in maintaining animal health by aiding the body in resisting the invasion and effects of external pathogens, thereby preventing wound infections (Esmaeili, 2021). In fish, this blood component is principally produced from head kidney (Kondera, 2011). Nevertheless, a portion of white blood cells remains present in the body’s tissues to fulfill the task of protection, identifying and neutralizing disease-causing agents. An elevation in the quantity of these cells often indicates the presence of infections stemming from bacteria, parasites, or other pathogens. Additionally, in certain instances, an increase in white blood cell count under normal condition may result from chronic diseases or abnormalities within the immune system. White blood cell indicators are an useful tool to diagnose fish health after supplementing with prebiotics or other products. In our study, we did not observe any increase in white blood cell indicators. This can be explained by the fact that peptidoglycan is only a component of the bacterial cell wall, primarily stimulating the humoral immune system rather than cellular immunity (Nguyen et al., 2020). Indeed, previous studies testing peptidoglycan supplementation in aquatic animal feed all noted an increase in humoral immune factors such as lysozyme and complement (Zhou et al., 2006).
Indeed, the influence of peptidoglycan supplementation in diets were recorded in immune variables measured in fish blood plasma. Two approaches are commonly employed to enhance the immune response of aquatic animals using prebiotics: one involves promoting the growth of gut microbiota, while the other entails activating the non-specific immune system (Song et al., 2014). Prebiotics can stimulate pro-inflammatory and complement responses; however, the effects of prebiotics are vastly different among fish species (Song et al., 2014). Since infection in defense mechanisms, the cumulated peroxidases contained in cells are released, causing an increase of level of both immune indicators at the initial time in plasma and progressively decreasing over time (Peixoto et al., 2019). Lysozyme is a bacteriolytic enzyme of the nonspecific system in most animals. The main sources of lysozyme are monocytes, macrophages and neutrophils. The bactericidal activity of this enzyme involves the hydrolysis of bacterial cell wall peptidoglycan produced during cell lysis (Nawaz et al., 2022). In our case, peptidoglycan is considered an exogenous substance from gram-positive bacteria (Lactobacillus sp.). Fish immune system recognizes and reacts to eliminate that substance out of the body (Lin et al., 2019), therefore, the lysozyme activity in PL10-fed fish was higher than PL0 group after 2 weeks of feeding trial. The lower dose of dietary peptidoglycan (PL2.5 and PL5) did not induce the increase of this immune parameter in striped catfish juveniles. On the other hand, the influence of dietary supplementation was observed after 4 weeks of nutritional experiment for peroxidase activity indicating that the effect of peptidoglycan supplementation was observed latter than lysozyme activity. This result may be explained by substrate competition between these two immune targets in early period of feeding trial (T2), the action of lysozyme then decreased and replaced by peroxidase activity after 4 weeks of experiment.
Based on the obtained results, we conclude that dietary supplementation induced the positive impacts on the feed utilization and humoral immune responses. However, in the normal rearing condition, the dietary supplementation at 2.5 g/kg of feed is suggested. The increases of immune variable in fish fed on the diet supplemented with peptidoglycan at 10 g/kg of feed suggest to test the effect of peptidoglycan on the disease resistance.