
Introduction
Labeobarbus intermedius (Rüppell, 1836) is an extensively distributed fish species in most parts of the Ethiopian catchments and Northern Kenya basins (Dadebo et al., 2013). It is widespread in the Ethiopian Rift valley, Abay, and Baro-Akobo basins of Ethiopia where Lake Tana hosts the largest population (Awoke, 2015; Vijverberg et al., 2012). However, despite the abundance and socioeconomic importance of L. intermedius in Lake Tana, the catches are now declining from time to time due to overfishing, habitat destruction, irrigation, and dam construction (de Graaf et al., 2000; Dejen et al., 2017; Gebremedhin et al., 2019; Mingist & Gebremedhin, 2016).
L. intermedius feeds on a variety of foods in various aquatic environments including zooplankton, phytoplankton, aquatic insects, fish, detritus, fish scales, nematodes, macrophytes, gastropods, fish eggs, fishes, mollusks, benthic invertebrates, insect larvae, and nematode (Dadebo et al., 2013; de Graaf et al., 2000; Deribe et al., 2011; Desta et al., 2006; Engdaw, 2014; Sibbing et al., 1994; Wakijira, 2013). Those studies also showed that there is a variation of food items within the same study area and fish species at different times of study. This is due to prey availability, season, habitat difference, and size of the fish (Desta et al., 2006; Sibbing & Nagelkerke, 2000).
Investigating the diet of fish species is a subject of ongoing research as it provides a basis for the developing of a successful management program for capture fisheries and aquaculture (Shalloof et al., 2009). In addition, the information on the natural foraging and feeding habits of fish species allows the identification of the trophic relationships present in the aquatic ecosystem and the identification of the food composition, structure, and stability of food webs in the ecosystem (Otieno et al., 2014). Such information is also important to guide the management of fish in the natural environment and within the controlled environment and the formulation of an acceptable diet for the fish in aquaculture (Adeyemi et al., 2009). Therefore, understanding its food and feeding habits is a key factor to successful and sustainable fish farming in a controlled environment (Shalloof & Khalifa, 2009).
The Ribb Reservoir fishery started in 2018 and L. intermedius is one of the commercially most important fish species in the Ribb Reservoir fishery (Mequanent et al., 2022). Despite their commercial importance, there is no information on the biology (food and feeding habits) of this species with the interaction of the ecology of the reservoirs. Exploring such type of information in Ribb Reservoir plays a fundamental role in the sustainable use of the stock and in improving the livelihoods of newly formed fish associations around the reservoir. Therefore, the present work aimed to elucidate seasonal and ontogenetic dietary changes and the feeding habits of L. intermedius species in the Ribb Reservoir.
Materials and Methods
Ribb Reservoir (11°59'0" N to 12°2'0" N latitude and 38°0'0"E to 38°2'59"E longitude) is an earthen rock dam with a capacity of 234 million m3 of water with a height and length of 73.5 m and 800 m, respectively (Ministry of Water Resources of Ethiopia, 2010) (Fig. 1). The reservoir dammed on the Ribb River which is the main tributary of Lake Tana and originates at a distance of 90 km from Gunna Mountain and bounded Farta, Ebinat, Libo-Kemkem, and Fogera districts of the South Gondar Zone, Amhara Regional State (Ashenafi, 2011). The reservoir is situated mainly in the Farta district with a small part engulfing the Ebinat district of the South Gondar Zone. It is found in the Lake Tana sub-basin, in the northern part of Ethiopia and the northeastern part of the Blue Nile basin and the sub-basin has an area of 15,320 km2 (Amhara Design and Supervision Works Enterprise (ADSWE), 2015). It is located at a distance of 625 km north of Addis Ababa and 60 km from Bahir Dar town, the capital city of the Amhara Regional State (Mulatu et al., 2018).

The boundary of the study area is classified as a tropical highland monsoon climate namely Wurch on Mount Gunna, Dega around Debre-Tabor town and Woina Dega near Ribb Reservoir (Ezezew, 2019). The climate of the Ribb basin was characterized by a rainy season (May to September) with 65 mm of monthly precipitation in May and 411 mm in July (Sisay, 2017). While the highest and lowest average annual rainfall of the basin were recorded as 1,400 mm and 1,200 mm, respectively (Bezabih, 2021). Whereas, there is no recorded rainfall data in the dry season. Also, the temperature of the Ribb basin varies throughout the year, from 19°C (December) to 23°C (May), and sometimes maximum temperature record of 30°C and the minimum temperature of 11.5°C (Bezabih, 2021).
L. intermedius samples were collected during the wet season (June–August) and dry season (December–March) using hooks and lines, and gillnets of stretched mesh sizes of 4, 6, 8, 10, and 12 cm. Total length, fork length (FL), and standard length were measured to the nearest 0.1 cm using the measuring board. Also, the total weight (TW) was measured to the nearest 0.1 g by using a sensitive balance. Then, the fish sample was dissected with a scissor and checked for gut fullness. The non-empty guts were preserved in a 5% formaldehyde solution for further analysis.
Identification of large foods was performed visually whereas smaller foods were identified by using a dissecting microscope (LEICA S4E, 10X/23) and compound microscope (OPTIKA E-PLAN, 10X/0.25). The relative importance of each food item to the diet of each fish species was estimated as follows.
The number of gut content samples in which a given food item containing one or more food items was expressed as the percentage of all non-empty guts (Bagenal & Braum, 1978). The proportion of the population that feeds on a particular food item was estimated by this method.
Food items that are found in the guts were sorted into different taxonomic groups and the volume of each group of food items was measured (Bowen, 1983). Then, the volume of a given category of food items was expressed as a percentage of all the categories of food items present in the gut samples. For macroscopic food items, the volume of the displaced water within the measured cylinder by the food items was expressed as the volumetric percentage of the measured food item as developed by Hyslop (1980).
The seasonal variations in the type of food eaten by L. intermedius were studied as a function of dry and wet seasons using a percentage of frequency of occurrence (%FO) and volumetric proportion. For the ontogenetic change in diet, the fish samples were determined by dividing their length into three size classes (I: < 20 cm; II: 20–29.9 cm; III: > 30 cm) (Dadebo et al., 2013). The percentage mean volume of various foods was calculated using the methods of Schoener (1970) and Wallace (1981).
A Chi-square (χ2) test was used to compare the frequency of occurrences of the different food categories during dry and wet season samples (Sokal & Rohlf, 1995). Whereas, the independent sample t-test was used to compare the volume of the different food categories consumed during wet and dry season samples. Dietary overlap between different size classes was calculated as percentage overlap using Schoener diet overlap index (SDOI) (Schoener, 1970; Wallace, 1981):
Where, α is the percentage overlap of the SDOI between size class ‘a’ and ‘b’. Pai and Pbi are the proportions of the food type i used by size class ‘a’ and ‘b’, and n is the total number of food types. When the α value exceeds 0.60 the diet overlap index is said to be biologically significant (Mathur, 1977).
Results
A total of 203 L. intermedius samples collected during the dry season (December to March) and wet season (June to August) were used for gut content analysis. The size of the fish samples ranged from 14.5 to 44 cm FL and weighed 56 g to 1,063 g TW. Of these, 71 (35%) were empty, while 132 (65%) contained food in the stomach. The diet of L. intermedius from Ribb Reservoir consists of macrophytes, phytoplankton, detritus, insects, zooplankton, nematodes, and unidentified materials. Of these foods, the most important foods were macrophytes, phytoplankton, detritus, and insects. While the remaining foods, zooplankton, nematodes, and unidentified materials, were relatively less important foods in their stomachs.
The %FO of macrophytes, phytoplankton, detritus, zooplankton, and unidentified materials in the diet of L. intermedius was significantly different between seasons (χ2 test, p < 0.05). Similarly, the volumetric contribution of macrophytes, phytoplankton, detritus, zooplankton, and nematodes differed significantly between seasons (χ2 test, p < 0.05) (Table 1). Whereas the %FO and the volumetric contribution of insects did not differ significantly between seasons. Macrophytes and detritus were the major dietary components in the diet of L. intermedius during the wet season, occurring in the gut at 63.5% and 71.4% in the gut and contributing 50.3% and 23.3%, respectively, to the total volume of dietary components (Table 1). During the dry season, the %FO of macrophytes (14.5%) and detritus (18.8%) was lower. Similarly, the contribution of both macrophytes (5.4%) and detritus (5%) decreased in the dry season. The volumetric contribution of phytoplankton was the first predominant food item in the diet of L. intermedius, occurring in 78.3% and accounting for 58.2% of the total volume of food items in the dry season. Zooplankton occurred in 23.8% and accounted for 4.8% of the total amount of food in the wet season. During the dry season, the abundance of zooplankton increased to 55.1% and contributed 13.7% to the total volume of food.
The %FO and volumetric contribution of insects in the dry and wet seasons did not vary significantly, occurring 56.5% in the dry and 58.7% in the wet season. And contributed to 15.2% and 12.2% in the dry and wet seasons, respectively (Table 1). From the insect groups, Diptera were the most predominant taxa compared to the other taxa, contributing 8% in the wet season and 6.7% in the dry season.
SDOI revealed a significant dietary variation between I & II (0.45) and I & III (0.30) size classes but not II & III (0.69) size classes (Table 2).
| Size classes | SDOI value |
|---|---|
| Lower (< 20 cm TL) and intermediate (20–30 cm TL) | 0.45* |
| Intermediate (20–30 cm TL) and higher (> 30 cm TL) | 0.69 |
| Lower (< 20 cm TL) and higher (> 30 cm TL) | 0.30* |
In the smallest size class < 20 cm FL, the mean percentage volume of food was dominated by insects (35.5%), nematodes (18.9%), zooplankton (18.6%) and detritus (12.6%). In contrast, the contribution of other foods such as phytoplankton (7.7%), macrophytes (5.7%) and unidentified materials (1.0%) was relatively small (Fig. 2).
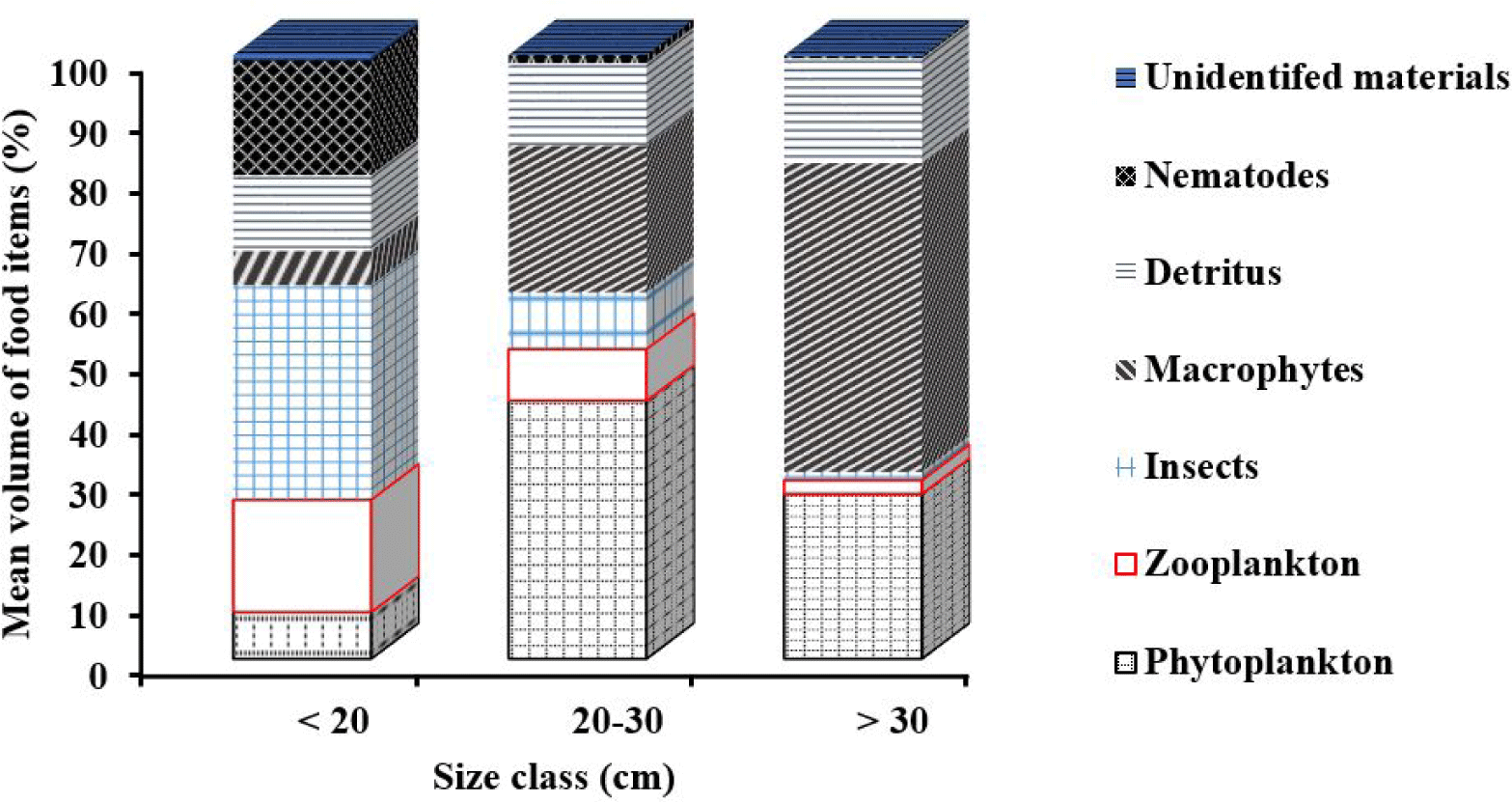
In the intermediate size class (20–30 cm FL), the volumetric contribution of phytoplankton (42.7%), macrophytes (24.3%) and detritus (13.7%) were higher than those of insects (9.4%), zooplankton (8.5%) and nematode (1.4%) (Fig. 2). In the higher size class > 30 cm FL, the importance of macrophytes and detritus further increased to 51.1% and 17.4%, respectively. On the other hand, the importance of zooplankton, insect, nematodes, and unidentified materials was almost negligible (Fig. 2).
Discussion
The present study demonstrated that L. intermedius fed on macrophytes, phytoplankton, detritus, insects, zooplankton, nematodes and unidentified materials in Ribb Reservoir. Of these foods, the first four accounted for the majority of foods (85%). While the remaining three made up 15% and are less important in the diet of L. intermedius (Table 3). Similar diet composition has been reported by different authors in different bodies of water such as Lake Tana (de Graaf et al., 2000; Engdaw, 2014); Lake Koka (Dadebo et al., 2013) and Tekeze Reservoir (Gebru, 2020). However, snails and fish scales have been reported in these waters, but not in the present study. In contrast to the present study results, the main food items of L. intermedius are mollusks (90%), fish prey and aquatic insects in Lake Awassa (Desta et al., 2006).
The most important foods in the diet of L. intermedius were insects, occurring in 57.6% of the intestines and 13.6% of the volumetric contributions. Among insect groups, Diptera was strongly dominant compared to other groups, its volumetric components were 7.4% (Table 3). This may be due to the dense stand of aquatic weeds supporting significant biofilm accumulation of benthic prey production as noted by Piet (1998). This finding is also consistent with Dadebo et al. (2013) in Lake Koka.
The contribution of food consumed by L. intermedius depends on the season, prey availability, habitat type and fish size (Desta et al., 2006). The African and Asian inland waters also showed that L. intermedius was polyphagous and can change its diet depending on seasonal and spatial differences in prey (Sibbing & Nagelkerke, 2000). Seasonal variations have a major impact on food availability and emergence (Desta et al., 2006). This is also underpinned by the current study result on the Ribb Reservoir. The feeding habits of L. intermedius in the present study are consistent with the omnivorous nature of L. intermedius studied elsewhere in the Gilgel-Gibe Reservoir in the OmoTurkana basin (Wakijira, 2013).
In the present study, L. intermedius showed significant seasonal variation in their feeding activity in Ribb Reservoir. This may be due to seasonal variations and the abundance of food in the reservoir. L. intermedius consumed phytoplankton mainly in the dry season and was almost absent in the wet season (Dadebo et al., 2013). This could be due to the high succession ability of phytoplankton during the dry season and the turbidity behavior of the reservoir in the rainy season (Dadebo et al., 2014; Wondie et al., 2007). In turbid waters, phytoplankton abundance decreased due to reduced light and low photosynthetic activity. All inflowing tributaries of the Ribb Reservoir bring a lot of sediment from the degraded catchment areas, resulting in turbid water in the reservoir, especially at the estuaries of the Ribb, Melo, Hamus Wonz and other rivers. Similar reports have been made in different water bodies of Ethiopia such as Lake Chamo (Fekadu & Chanie, 2017). In addition, Wondie et al. (2007) also reported that phytoplankton biomass decreased during the rainy season as a result of the increased turbidity of Lake Tana.
During the wet season, macrophytes and detritus were the most important food items in the diet of L. intermedius, this result agrees with the study of Wakijira (2013) in Gilgel Gibe Reservoir and Dadebo et al. (2013) in Lake Koka. It was reported that macrophytes and detritus are the most important food items during the wet season and the source of the detritus is likely to be the floods that brought fragments of plant matter (Dadebo et al., 2013, 2014). In this study, the contribution of insects to the diet of L. intermedius was comparable in both the dry and wet seasons. However, some taxa contributions varied in both the dry and wet seasons. In general, seasonality affects the feeding behavior and prey organism species of L. intermedius in the Ribb Reservoir. During the rainy season, the contribution of foods of plant origin was higher than the contribution of foods of animal origin.
The SDOI rule is evident that L. intermedius showed a slight ontogenetic dietary shift in the Ribb Reservoir. There was a significant ontogenetic dietary shift between size classes I and II (= 0.45) and between size classes I and III (= 0.30). However, there is no significant ontogenetic dietary shift between size classes II and III (= 0.69). These non-significant results between these size classes of fish could be due to the development of similar morphological and physiological traits and the feeding of similar foods. This is because the ontogenetic dietary transition in fish is determined by the morphological and physiological characteristics of the fish (Agembe et al., 2019).
The diet composition of L. intermedius showed that the foods in each size class were digested, but at different rates. For example, insects, zooplankton, and nematodes have been heavily included in smaller size classes, but decreased with the size of the fish. On the other hand, the uptake rate of macrophytes and detritus increased with the size of the fish. This size-dependent dietary change may be due to the morphological and physiological development, water type, and habitat of the fish (Engdaw et al., 2013). Most juveniles prefer to swim in the coastal region, feeding on the invertebrate molluscs and zooplankton that are prevalent there (Dadebo, 2009). As they grow in size, they swim in open water and feed on large dietary components of macrophytes, detritus, and others related to the morphology and physiological traits of the fish.
In this study, the composition of insects and macrophytes (the most important foods) showed significant differences in different size classes. For example, insects were consumed heavily in smaller classes but less in larger classes. Whereas macrophytes were heavily consumed by the larger size class, while consumed less in the lower size class. This study result agrees with the work of Desta et al. (2006) in Lake Hawassa, who found a slight ontogenetic dietary change. They found that insects are the most important food elements for juvenile fish, but their importance decreases with the size of the fish and shifts to piscivorous diets with increasing size, but not the fish prey observed in the current investigation. Furthermore, it has been reported that slight ontogenetic variation is observed in Lake Koka, where macrophytes are dominant foods in the higher-size class but are relatively less important in the smaller-size class diet (Dadebo et al., 2013). In this study, insects were the dominant foods in the smaller size class, but relatively less important in the larger size class.
In conclusion, this study has demonstrated the diet, seasonal variations, and ontogenetic dietary shifts of L. intermedius in the Ribb Reservoir. The main components of the diet were macrophytes, phytoplankton, detritus and insects. While the rest of the foods in his diet were less important. The present study also showed seasonal variations and dietary changes in the size of the fish. In the dry season, zooplankton and phytoplankton were the main diets, while in the wet season, macrophytes and detritus were the predominant diets. The SDOI rule shows that there was significant diet variation of size class I compared to size classes II and III of L. intermedius and a slight ontogenetic dietary shift ensued. However, size classes II and III did not show any significant change in diet.