





Introduction
Shark species within the Lamnidae family exhibit heightened vulnerability compared to other shark species, attributed to various biological traits, including slow growth, a protracted reproductive cycle, and low fecundity (Brooks et al., 2010; Campana et al., 2005; Semba et al., 2011). The 18th Conference of the Parties of the Convention on International Trade in Endangered Species of Wild Fauna and Flora (CITES) in September 2019 added two mako sharks species to Appendix II, which is longfin mako sharks (Isurus paucus) and shortfin mako shark (Isurus oxyrinchus) (Sellheim, 2020). The later also designated as an endangered species by the International Union for the Conservation of Nature and Natural Resources. The main reason was the rapid declines above 96% in the Mediterranean, with projected of 60% in the North Atlantic and 41.6% in the Indian Ocean over the next decade (CITES, 2019).
Shortfin mako sharks is categorized as active and fast swimmers species that migrate widely in tropical and temperate waters above 16°C. (Barreto et al., 2016; Compagno, 2001). As a fisheries commodity with a high economic value, shortfin mako shark particularly vulnerable to overfishing (Ferretti et al., 2008; Musick et al., 2000). Moreover, the shortfin mako shark is a bycatch of fishing methods such as long lines (Sulaiman et al., 2018; Wu et al., 2021) and gillnet in Indonesia (Prihatiningsih et al., 2018). The overfishing condition, slow growth, and low fecundity caused the mako shark to be highly vulnerable to stock depletion, resulting in massive economic losses in the fisheries industries.
Biological characteristics (including length of maturity, asymptotic length, and coefficient of growth) and stock status information are crucial for managing shark fisheries (Yokoi et al., 2017). The stock status required growth information that can be estimated using aging through the hard part of fish such as otolith, vertebral, or spine. However, using these methods is expensive. Alternatively, length (size) data can be used to estimate the growth parameter and predict stock status. Some information on the age and growth of mako sharks have been reported using vertebral analysis, revealing a slow growth and needing a long period to reach maturity (Groeneveld et al., 2014; Semba et al., 2009). Moreover, some previous studies have reported the overfishing condition for some species of sharks in the Indian Ocean, such as a study on Sphyrna lewini using four years of size data from 2013 to 2016, Alopias pelagicus using two years of size data from 2015 to 2016 and Prionace glauca using two years of size data from 2019 to 2020 (Chodrijah et al., 2021; Chodrijah & Faizah, 2021a, 2021b).
The latest study with multiple years of size data is essential to evaluate the stock status of shark fisheries in the Indian Ocean, especially in the south of Java and West Nusa Tenggara waters. A length-based spawning potential ratio (SPR) can be used to study the stock status and to investigate an optimal fishing level for the exploited stock (Hordyk et al., 2016). This study used a length-based assessment to evaluate the stock status of I. oxyrinchus by using eight years of data to develop a management strategy for shark fisheries. Hopefully, the findings will contribute to sustainable fishing practices, ensuring the preservation of shark populations and the long-term health of the marine ecosystem.
Materials and Methods
A daily scientific monitoring program was conducted from mid-2013 to 2021 (except for 2017) in the Cilacap Ocean Fishing Port, Central Java, and Tanjung Luar landing site, West Nusa Tenggara to collect biological information on the shortfin mako shark (I. oxyrinchus) (Fig. 1). Each shark landed was measured on-site as total length (TL) to the nearest lower cm, weighed as total weight in kg, maturity stage recorded, and sex based on the clasper presence. There was 1,386 length frequency information obtained during the study, comprised of 859 females and 527 males.
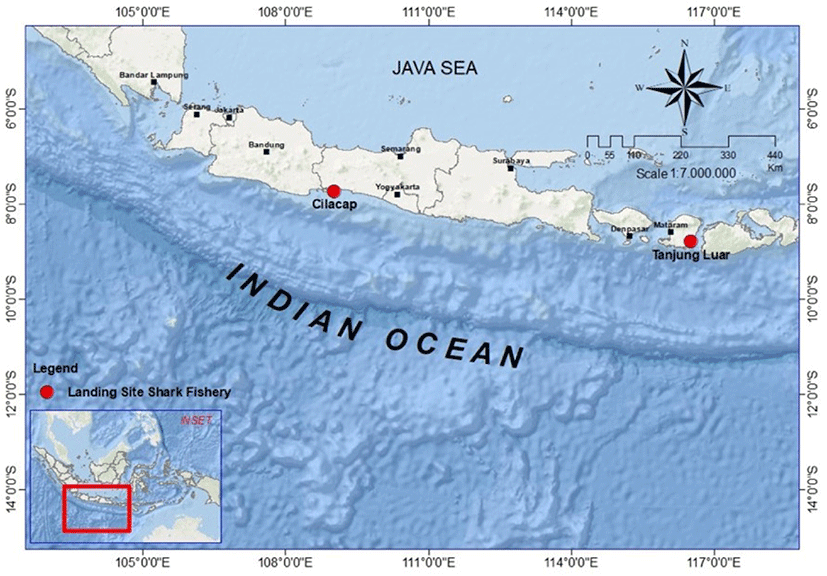
The annual sex ratio of I. oxyrinchus was analyzed with the chi-square test to identify the number between males and females. A histogram was performed on the annual TL frequency data. The length at first maturity followed White (2007) was 185 cm TL for male. As for females, Lm50 was at 248 cm.
The growth of I. oxyrinchus can exhibit seasonal fluctuations caused by environmental factors, including variations in water temperature throughout different seasons. To account for this seasonal pattern, the standard von Bertalanffy growth function (VBGF) can be modified by introducing a seasonal oscillation as described by (Somers et al., 1987):
Lt is the total length at age t; L∞ is asymptotic length; K is the growth coefficient; t0 is theoretical age at zero length.
Since I. oxyrinchus do not change sex during their life cycle and exhibit sexual dimorphism in size, indicating distinguishable growth patterns (Tsai et al., 2014), the seasonal VBGF were applied to males and females separately using the electronic length frequency analysis (ELEFAN) in TropFishR packages from the R version 4.2 (Mildenberger et al., 2017).
The theoretical age at length zero (t0) of I. oxyrinchus was calculated using Pauly (1983) equation:
The asymptotic length (L∞) and growth rate (K) were used to estimate the natural mortality of I. oxyrinchus. The natural mortality was estimated based on the equation (Then et al., 2015):
Moving average (MA) and the optimum number of bins (OBS) settings can significantly affect the scoring of the growth curve (Mildenberger et al., 2017). Therefore, the calculation of OBS follows Wang et al. (2020), where OBS=0.23 × Lmax0.6, resulting in 7 and 8-bin for males and females, respectively. As for MA, aside from the default setting in FISAT II (Gayanilo et al., 2005), i.e., a width of 5 bins. The other scenarios were also tested as sensitivity, with 7, 9, and 11-bin cohort widths.
The length-based SPR is one of various tools for investigating the status of fisheries stock. The SPR of I. oxyrinchus in this study was estimated using the LBSPR package in the R program with some input parameters: length-frequency data, M/K ratio, and length at maturity (Hordyk et al., 2016; Mildenberger et al., 2017; R Core Team, 2023). The length at selectivity and F/M were measured based on the outputs of the length-based spawning potential ratio (LB-SPR) packages in the R program. The 40% SPR was used as a reference point for the optimal stock of the shortfin mako shark, borrowing from Northwest Atlantic (Tsai & Huang, 2022).
Results
The sex ratio between males and females of I. oxyrinchus was 1:1.63, indicating that the proportion of female shortfin mako sharks was significantly higher than that of males. The pattern happened throughout the years of observation, except for 2013. The size ranged from 25–363 cm TL for females and 69–310 cm TL for males, with the mean size around 192.35 ± 41.61 cm TL and 195.72 ± 35.72 cm TL (Table 1).
Referring to the study of White (2007), where males matured at 185 cm TL and females at 250 cm, therefore, almost all female specimens (92%) were found to be immature, while only around one-third of the males sampled (34%) were already mature (Fig. 2).
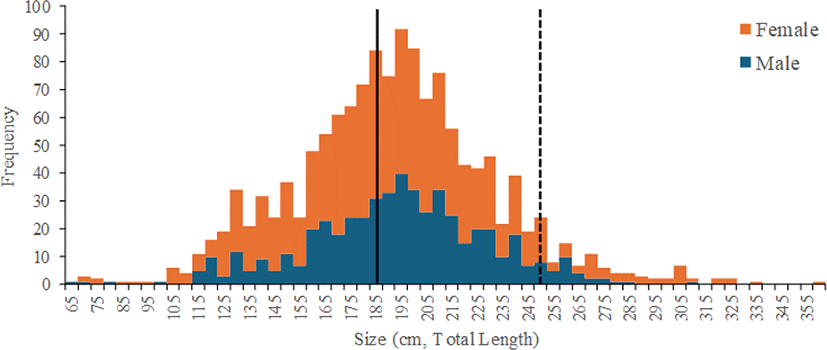
MA did affect the estimation of asymptotic length (L∞) and growth coefficient (K) under various sexes (male, female, and pooled) and methods scenarios (genetic algorithm [GA] and simulated annealing [SA]). Generally, the lower the MA, the higher the goodness of fit (Rn max) value. Based on the highest Rn max value, MA = 5 best fit this species. However, between the two methods, SA gave a higher value of L∞ (~50 cm TL) but a lower K (~0.01) than the GA. Using SA, the growth rates (K) ranged from 0.0368 to 0.1196 /year. Using SA, the growth rates (K) ranged from 0.0368 to 0.1196 /year. GA gives a higher growth rate, implying more rapid growth and a shorter lifespan than SA. Since SA tended to produce larger uncertainties, thus, GA was picked as the best option to analyze this species because it created a more plausible growth parameter. The L∞ and the K of I. oxyrinchus for females, males, and pooled sex based on the highest Rn value were 392.54, 344.34, 386.86 cm TL and 0.0368, 0.0477, 0.0396 /year, respectively (Table 2).
The theoretical age (t0), when the shark has zero length, was –4.02, –3.19, and –3.75 years for females, males, and pooled sex, respectively. Male sharks attained their length at first maturity (Lm50 = 185 cm) at 14 years, and female sharks attained their length at first maturity (Lm50 = 248 cm) at 23 years (Fig. 3). The maximum age of males and females sampled in this study was estimated up to 66 and 45 years. The natural mortality (M) for females, males, and combined sexes were estimated at around 0.0524, 0.066, and 0.0555 /year, respectively.
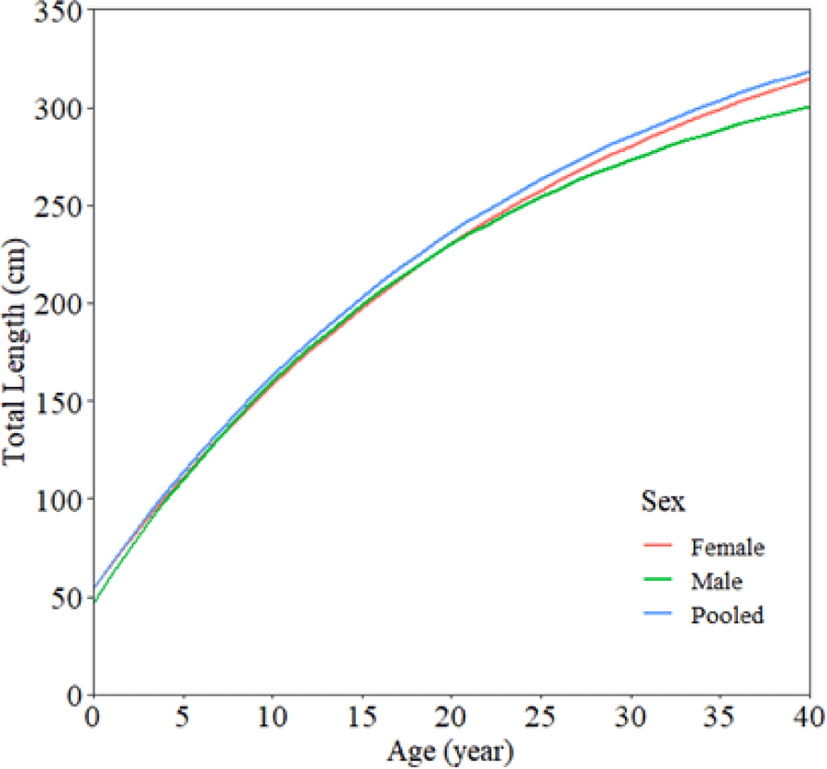
The length-based SPR used some inputs parameters, including the length-frequency data, asymptotic length (L∞) as 386.86 cm TL, the fishing mortality and natural mortality ratio (M/K) as 1.4, length at 50% maturity (Lm50) as 248 cm TL and length at 95% maturity (Lm95) as 370 cm TL. The selectivity (SL50) of I. oxyrinchus caught by longline in the Indian Ocean gradually decreased from 181.78 to 160.84 (Fig. 4). The F/M ratio was much higher than one, ranging from 2.48 to 4.53. The yearly SPRs from 2013 to 2021 are around 4% to 8%, with the SPR of 6% in 2021.
Some uncertainties in the SPRs were driven by the variation of some life history parameters, including asymptotic length (L∞), growth rate (K) and natural mortality (M). The lower value of asymptotic length will impact the higher estimates of the SPRs. Among all the variations of asymptotic lengths and growth rates, the SPRs from 2013 to 2021 ranged from 0.02 to 0.14 (Table 3). The SPR of I. oxyrinchus in 2021 was 0.05 ± 0.02 (mean ± SD).
Discussion
The size distribution of shortfin mako shark in the Indian Ocean south of Java, Bali, and Nusa Tenggara ranged from 69 to 363 cm TL which is relatively similar to other areas reported worldwide. Barreto et al. (2016) found the size of the mako shark ranging from 73 to 296 cm TL, captured by longlines in Brazilian waters. However, Varghese et al. (2017) found a smaller size ranging from 70 to 269 cm TL in the Eastern Arab Sea. Interestingly, this study found that the mean size of female I. oxyrinchus tends to decrease along observation time, similar to what Roff et al. (2018) reported in Australia. A declining size in some commercial fisheries can be the impact of overfishing and low gear selectivity, as is the case for reef fishes in the Java Sea (Tirtadanu et al., 2023). Continuously high fishing pressure in a proximity area could cause local stock depletion of this species, resulting in a shrinking size.
The proportion of female I. oxyrinchus was almost twice as much as the male, which indicated an unbalanced sex ratio. The overall sex ratio of shortfin mako shark in the Moroccan Central Atlantic Coast tends towards equilibrium (Alahyene et al., 2022). Chodrijah et al. (2021) also found a similar situation for thresher sharks in the same fishing ground. The dominance of female shortfin mako sharks around Nusa Tenggara suggests a potential spawning ground for pelagic sharks. However, further research is necessary to confirm this hypothesis. In general, the unbalanced sex ratio could increase sharks’ vulnerability. The high fishing pressure on the female shark can obstruct the spawning biomass and stock-recruitment process.
Some studies have reported the growth parameters of I. oxyrinchus in some areas, with the asymptotic length ranging from 285 to 387 cm TL and the growth rate (K) ranging from 0.01 to 0.16 /year (Table 4). Previous studies have mentioned an indication of sexual dimorphism in I. oxyrinchus, where females tend to have a larger size than males (Groeneveld et al., 2014; White, 2007). This study also found similar conditions where the size of females (L∞ = 392.54) was larger than males (L∞ = 344.34). The estimated asymptotic length (L∞) in this study was higher than in the Pacific and Atlantic Oceans. However, the growth coefficient (K) was relatively similar to vertebrae studies. Shortfin mako shark has a large body size and slow growth that increase the vulnerability of this species to overfishing (Cortés et al., 2010). Moreover, the shortfin mako shark needs a lot of time to reach the length at first maturity. Thus, immature specimens dominated the samples.
| Area | Length (cm ) | L∞ (cm) | K (/year) | Source | Methods |
|---|---|---|---|---|---|
| North Atlantic | 253–366 (FL) | 366 (FL) | 0.08–0.12 | Natanson et al. (2006) | Vertebrae |
| Western Atlantic | 73–296 (FL) | 340 (FL) | 0.04–0.13 | Barreto et al. (2016) | Vertebrae |
| Central Atlantic | 60–317 (TL) | 313 (TL) | 0.15 | Alahyene et al. (2021) | Length frequency |
| North Pacific | 73–330 (PCL) | 340 (PCL) | 0.09–0.16 | Semba et al. (2009) | Vertebrae |
| Pacific, Chile | 69–300 (TL) | 296 (TL) | 0.07–0.08 | Cerna & Licandeo (2009) | Vertebrae |
| New Zealand | 100–347 (FL) | 302 (FL) | 0.01–0.05 | Bishop et al. (2006) | Vertebrae |
| Southwest Indian Ocean | 91–311 (FL) | 285 (FL) | 0.11 | Groeneveld et al. (2014) | Vertebrae |
| Indian Ocean, South Java | 69–325 (TL) | 387 (TL) | 0.04 | Present study | Length frequency |
The suspicion of overfishing could be seen in the F/M ratio, which, in this study, was higher than the optimal condition. Gulland (1983) suggested equal fishing and natural mortality values (F/M = 1) as the threshold for optimal fisheries conditions. Overfishing was the main factor resulting in the depletion of sharks globally, driven by the high demand and the high economic values of shark fins (Dulvy et al., 2021). Moreover, the evidence of the declining mean body size of the female I. oxyrinchus from this study can be the impact of the overfishing condition on shark fisheries. The continuing shark fishing condition will increase the risk of the mako shark extinction in the Indian Ocean, southern Java.
The high-fishing pressure of shark fisheries in the Indian Ocean has caused low spawning stock biomass, called recruitment overfishing conditions. It was indicated from the SPR in 2021 (SPRcur = 6%), which is less than the target reference of 40%. Moreover, the high catch intensity of immature sharks (indicated by low SPR value) was also reported in some areas in Indonesian waters: S. lewini in Lampung was 16%, and Carcharinus sealei in the Arafura Sea was 23% (Nugraha et al., 2023; Nurdin et al., 2023). If the fishing pressure continues at its current state, it will collapse shark stocks in the future.
Some management measures are critical to maintaining the sustainability of shark fisheries. Tuna RMFOs have banned shark fishing and shark finning with overfishing status, while CITES limit shark trading of particular species categorized in Appendix II. Porcher & Darvell (2022) suggested the protection of shark and their habitats, followed by the international agreement on their conservation. However, defining a dedicated marine protected area for highly migratory species like mako sharks will be challenging since the area has been a traditional shark fishing ground for artisanal longliners. While applying a minimum catch size might not be an effective management measure, a strict catch/trading quota (TAC) could be effective as long as it is followed by cooperative management among governments, fishers, traders, shark industries, and the local community which aimed to maintain its sustainability.
The current regulation of shark fisheries management is related to the I. oxyrinchus fishery TAC (Total Allowable Catch) of 320 fish/year for West Nusa Tenggara province with a minimum landing size of 250 cm based on Ministerial Regulation No. 21/2021. In fact, with this management, I. oxyrinchus is still in an overfished recruitment condition with an SPR of only 6%. By comparing the current SPR of 6% with the reference 40% SPR, reducing the TAC of I. oxyrinchus to 48 fish/year is necessary. In addition, the policy must be implemented successfully and MCS (Monitoring, Controlling and Surveillance) system is also critical to prevent IUU fishing in shark fisheries. Institutional strengthening and government regulations related to shark conservation must be well-socialized in the community (Sulaiman & Triharyuni, 2021). Simmons & Mehmet (2018) also suggested listening local communities and adopting policy communication in developing sustainable shark management. To maintain the economy of traditional fishers, governments must provide other job alternatives to replace mako shark fisheries. Strong integration between the government, local communities and the shark industry is critical to successfully implementing sustainable shark management policies.
The length-based approach method has its limitations. Samples obtained from two landing sites, representing Industrial (Cilacap) and artisanal (Tanjung Luar), are mainly from longline gear, potentially introducing a selectivity bias (Punt et al., 2014), resulting in a dominance of immature fishes in the samples. Broader sampling across various sites could improve assessment accuracy and capture population dynamics more effectively, providing a better representation. Kell et al. (2022) suggested that length-based indicators are more effective for long-lived species with slow growth rates, which is in line with species of interest in this study. They perform better when the focus is on protecting mature fish rather than immature ones, and they are more adept at indicating trends than quantifying harvest levels, even amidst uncertainty about population dynamics over time. Another limitation is that the length-based approach assumes the stock should be in equilibrium. Moreover, the growth rate and natural mortality should be constant (Chong et al., 2019). Therefore, the uncertainty of the spawning stock biomass (SSB) is crucial to investigate the sensitivity and to avoid the over-estimate of SSB, which leads to mismanagement. This study used some variations of asymptotic length, growth rate and natural mortality to understand the sensitivity of the SPR. Among all the variations of those life history parameters, the SPRs from 2013 to 2021 ranged from 0.02 to 0.14, with the mean SPR in 2021 being 0.05 ± 0.02 (mean ± SD). However, all of the estimations among the scenarios were much lower than the optimal condition of 40% SPR, indicating a high recruitment overfishing condition. If the fishing effort of I. oxyrinchus continues at this current level, the stock will collapse due to the collapse of the spawning stock biomass, with no recruitment. Some management effort options are crucial to maintaining sustainability, e.g., fishing gear regulation, critical habitat protection, fishing location regulation of fishing season and awareness raising. Furthermore, future studies must highlight specific management recommendations for optimal catch limit, sustainable fishing gear modification, and closure regulations based on area and season.