






Introduction
Indo-Pacific groupers (Family Epinephelidae) are high-level or top predators playing important roles in tropical coastal ecosystems (Sadovy de Mitcheson et al., 2013). These valuable food fishes are targeted to supply export and domestic markets across all or most of their ranges (Sadovy de Mitcheson et al., 2013, 2020). Epinephelid groupers are typically caught in multi-species fisheries, using a variety of legal and illegal gears (Halim et al., 2020; Khasanah et al., 2020). In Indonesian deep-water demersal fisheries, typically export-oriented and operating at depths of 50–500 m, groupers are often caught together with snappers (Dimarchopoulou et al., 2021). Meanwhile small-scale fisheries operating in shallower waters mainly on or close to coral reefs typically catch a wide range of coral-associated fishes, including groupers, with smaller or less valuable fish often consumed by the fishers or sold on local markets (Ambo-Rappe & Moore, 2019; Nadiarti et al., 2021). Many multispecies reef fisheries have been reported as unsustainable (Newton et al., 2007). Furthermore, there are strong indications of shifting baselines (sensuPauly, 1995) in the perception of fish abundance and/or size, in particular for groupers (Ainsworth et al., 2008; Sadovy de Mitcheson et al., 2020).
Grouper biodiversity and stock abundance are clearly important to maintain ecological, economic and food security roles in coastal and small island ecosystems (Sadovy de Mitcheson et al., 2013, 2020). Therefore, sustainable management of grouper fisheries will contribute to achieving the sustainable development goals (SDGs) set out in the 2030 Agenda for Sustainable Development, especially SDG 14 (Life Under Water) but also SDG 1 (No Poverty), SDG 2 (No Hunger), and SDG 12 (Responsible Consumption and Production).
Widespread destructive fishing and/or overfishing of grouper stocks are reported from many Indo-Pacific countries (Amorim et al., 2019; Sadovy de Mitcheson et al., 2013), including India (Chatla & Pamulapati, 2020) and Indonesia (Ernawati et al., 2021; Halim et al., 2020; Khasanah et al., 2020). Although a revised assessment based on the International Union for Conservation of Nature (IUCN) Red List criteria in 2018 classified 11.4% of groupers as being at risk of extinction, perhaps surprisingly in view of the generally high levels of exploitation, the revised conservation status of most grouper species was least concern (LC) (Sadovy de Mitcheson et al., 2020). However, the detailed assessments reveal a lack of data on key indicators (e.g., population trend listed as “unknown”) for species no longer listed as data deficient (DD), e.g., Epinephelus sexfasciatus (Craig, 2018).
One challenge in estimating the status of particular species or stocks is that, while fisheries catch (production) data are collected by national agencies in most countries, these data are rarely at the species level for tropical demersal species, especially reef-associated fishes such as groupers (Amorim et al., 2019; Chatla & Pamulapati, 2020). In Indonesia groupers are targeted in all 11 Indonesian Fisheries Management Areas (FMAs) (Dimarchopoulou et al., 2021). At fish landing sites and in fisheries statistics, groupers are generally recorded under the generic term of kerapu, which comprises most grouper genera, although in some cases one or more high-value species are recorded separately, while Plectropomus spp. are commonly referred to and/or recorded as sunu (Khasanah et al., 2020).
Research on grouper biology and population status in Indonesian waters has tended to focus on one or more (typically high-value) species (Achmad et al., 2020; Ernawati et al., 2021; Halim et al., 2020). While most studies have focused on relatively small geographic areas, a recent nation-wide multi-year study using innovative participatory methods and supported by The Nature Conservancy (TNC) has provided valuable data with a species-level resolution on deep-water grouper and snapper fisheries operating at depths of 50–500 m (Dimarchopoulou et al., 2021; Mous et al., 2019a, 2019b). Focused mainly on fisheries targeting export markets, the TNC study found widespread signs of overexploitation, even though mature individuals dominated the sampled catches of most species. Meanwhile, an ecosystems approach to fisheries management assessment of grouper and snapper fisheries in East Kalimantan province, FMA 713 (Nadiarti et al., 2021) found that juvenile groupers of some species were much more common on local markets than would be expected based on the deep-water grouper and snapper survey results. They also found indications that juvenile groupers were caught (as targets or by-catch) in fisheries operating in shallow-water (less than 50 m depth) coastal fisheries mainly targeting other species (and therefore not covered by the TNC study on deep-water grouper and snapper fisheries); in addition to sale on local markets, juveniles were supplied live to grow-out operators, a form of capture-based aquaculture that can threaten wild stocks (Sadovy de Mitcheson et al., 2013, 2020). Juveniles comprised the majority of catches from 2016–2017 in FMA 713 for 9 out of 10 grouper species monitored (Khasanah et al., 2020). Several species-specific studies also report declines over time in grouper abundance and/or size, including in protected areas such as the Komodo National Park (Pet et al., 2005).
The speciose genus Epinephelus typically contributes a high proportion of groupers sold on local markets in various regions of the Indo Pacific including India (Chatla & Pamulapati, 2020) and Indonesia (Jefri et al., 2015). However, compared to groupers exported for the life reef food fish trade (LRFFT) or as fresh/frozen commodities, little is known about the species and size distribution of the groupers caught, sold and consumed locally in Indonesia. This study aimed to provide data on the species composition and size/life-stage structure of Epinephelus spp. groupers caught and sold locally in the Indonesian province of South Sulawesi, most of which come from the catches of small-scale fishers operating in coastal or small island coral reef ecosystems. These data can inform management policies and contribute towards species-level status assessments.
Materials and Methods
The study was conducted from June to October 2022 in two cities (Makassar and Palopo) and ten regencies (Barru; Pangkajene Kepulauan, henceforth called Pangkep; Maros; Takalar; Jeneponto; Bantaeng; Selayar; Sinjai; Bone; and Luwu) that were known to be major fish landing sites for groupers caught by small-scale fishers in South Sulawesi (Fig. 1 and Table 1). These sites represent four fishing bases for the grouper fishing grounds in each of the three seaways (Makassar Strait, Gulf of Bone, and Flores Sea) around South Sulawesi. A detailed description of the study area is given in Kadir et al. (2022).

Data were collected at the main fishing ports used by small-scale fishers and selling to local markets or directly at local markets in each of the 12 city/regency areas (sites). All groupers from the genus of Epinephelus were photographed on site with permission from the fish mongers and other traders. The standard and total lengths (TL) of each Epinephelus specimen were obtained using the Rapid Scaling on Object (RASIO) method developed at Universitas Hasanuddin by A. A. A. Husain. The photographic method was used to enable a rapid and minimally invasive data collection acceptable to the fishers, fishmongers and traders who would not allow enumerators to measure all the fish using classical methods such as fish rulers or tape measures. Validation of the method used was performed on a sub-sample of the data. The principle of this method is scaling according to the ratio between an object of known length and the object of interest. In this case the fish specimen was the object of interest, and the measured reference object was a whiteboard marker or pen (typically 14.5 cm or 15 cm). A photograph of each fish was taken with a measured object placed beside the body of the fish. The camera window frame was positioned perpendicular to the fish specimen and a close-up photograph taken, ensuring the whole body of the fish (from the tip of the snout to the tip of tail) was included in the frame, as shown in Fig. 2. The species was determined based on these photographs with the aid of references including Allen et al. (2003) and Heemstra & Randall (1993), and on-line databases including FishBase (Froese & Pauly, 2022). The current taxonomic nomenclature (valid name) for each species was verified in the World Register of Marine Species (WoRMS) on-line database.

Each image was then copied to an excel worksheet where a line was drawn across the image along the measured non-fish object; the length of this reference line was recorded as Sg. The conversion ratio (R) was then obtained through dividing Sg by the actual length of the object (marker/pen) used, noted as Sa. Another line was then drawn across the image, from the tip of the snout to the tip of the tail and the length recorded as Ig. The total length of the fish specimen was then calculated through multiplying Ig by R. All measurements were made in cm, with a precision of 0.01 cm.
The data on species composition were tabulated by site in Microsoft Excel 365 and analysed by site and seaway using the on-line ranacapa suite (Kandlikar et al., 2018) which gives the observed and Shannon-Weiner alpha biodiversity indices, the Curtis-Bray and Jacquard beta diversity indices, a cluster analysis dendrogram, and a heatmap of species distribution. Statistical significance was evaluated at the 95% confidence level, using analysis of variance (ANOVA) for alpha diversity and multivariate analysis of variance (MANOVA) for beta diversity.
All total length data obtained for each species were used to estimate the life stage of each individual fish. The life stage determination method used was based on several references (Gullström et al., 2002; Nadiarti et al., 2015; Nagelkerken & van der Velde, 2002). The size range was divided into three and fish with TL < 1/3 maximum length were categorized as juveniles, those with TL between 1/3 and 2/3 of the maximum length as subadults, and those with TL > 2/3 maximum length as adults. Life stage data were then aggregated and tabulated by species and site for each sampling site in Microsoft Excel 36. Specific or unusual characteristics of the specimens recorded were noted, for example the presence of ripe eggs oozing from the genital pores of visibly gravid females. The species composition and size/life-stage data were compared with data on Epinephelus sp. catches from other grouper fisheries in Indonesia and some nearby countries.
Results
The total number of grouper specimens recorded was 3,398, representing 23 species in the genus Epinephelus. The summary by seaway (Table 2) and the heatmap showing the distribution and relative abundance of each species between the 12 sites (Fig. 3) show that the species caught and their relative contribution to catch volume varied between sites and seaways. Four species each contributed more than 10% to the total sample (Epinephelus areolatus, Epinephelus ongus, E. quoyanus, and Epinephelus fasciatus), collectively comprising over two thirds (69%) of the catch. Two species each contributed 5%–10% of the total catch (Epinephelus coioides and Epinephelus faveatus), while four species contributed 1%–5% each (E. sexfasciatus, Epinephelus maculatus, Epinephelus bleekeri, and Epinephelus corallicola). The remaining 13 species each contributed less than 1% to the total sample, collectively contributing less than 5%.

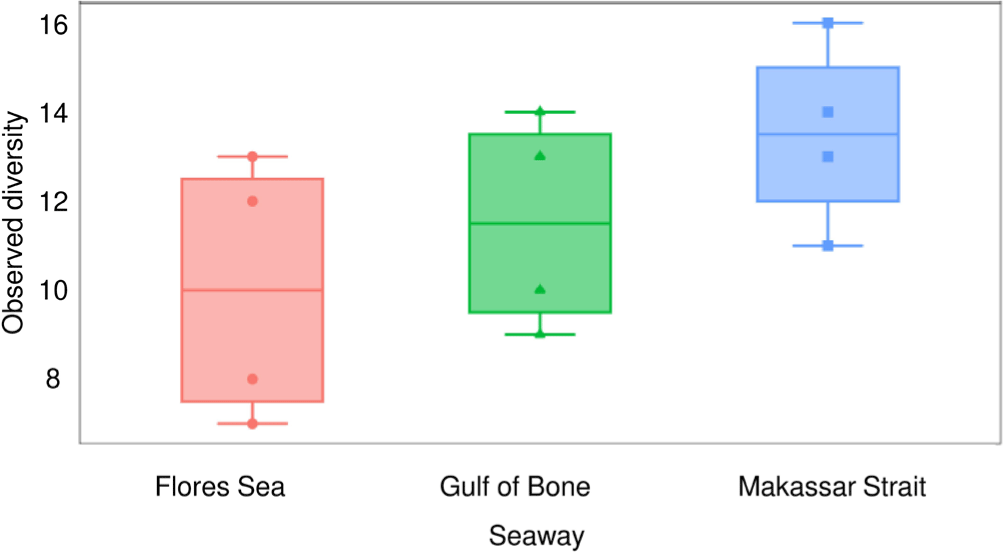
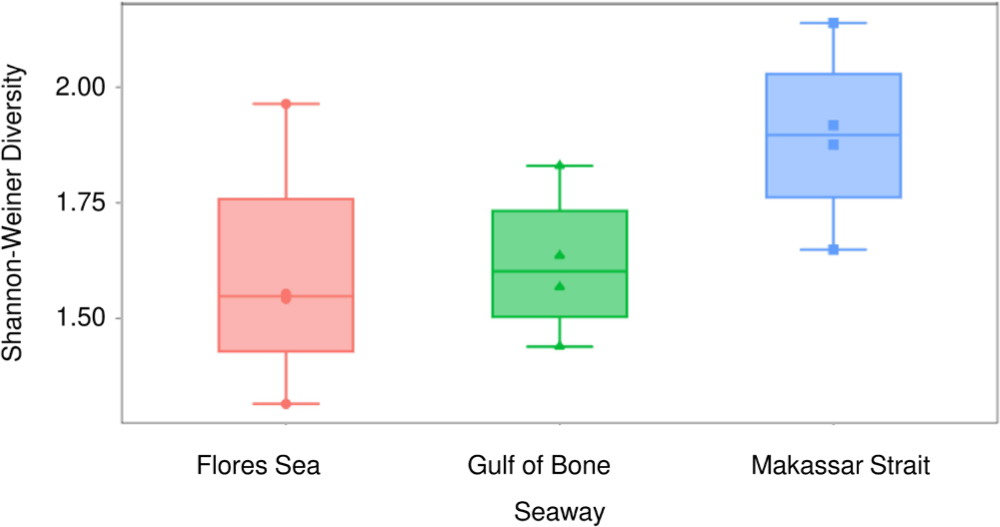
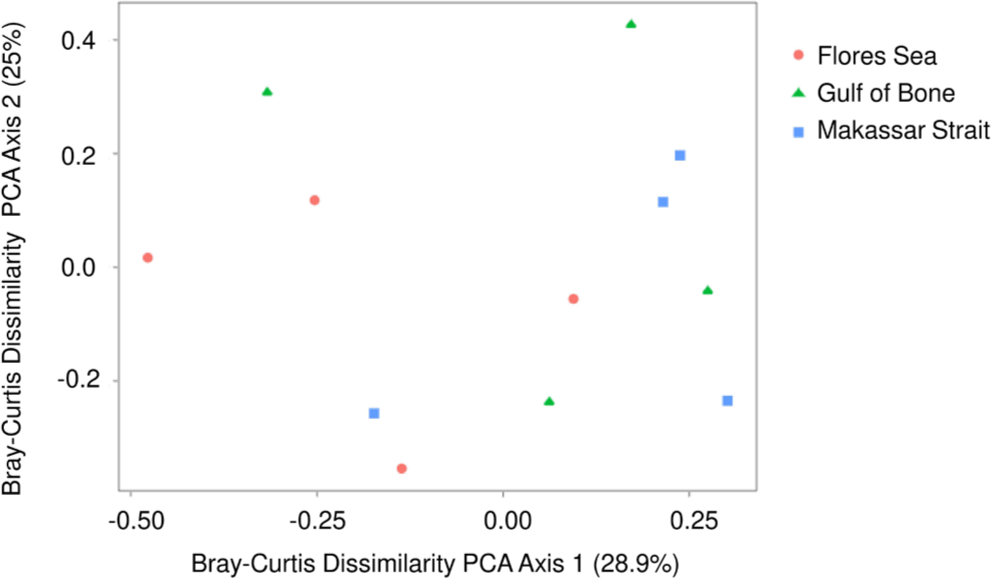
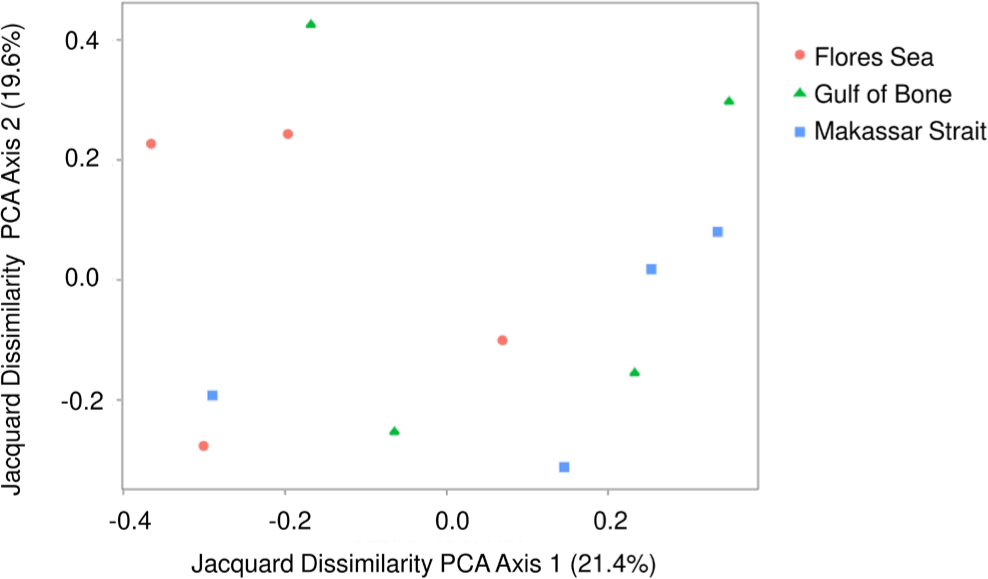
The alpha diversity plots appear to show considerable differences at the seaway level. Median values of the observed (Fig. 4) and Shannon-Weiner (Fig. 5) indices were lowest (respectively 10 and 1.55) in the Flores Sea, followed by the Gulf of Bone (respectively 11.5 and 1.60), and highest (respectively 13.5 and 1.90) in the Makassar Strait. However, the spread within each seaway was high and the differences were not statistically significant (ANOVA p > 0.05). The beta diversity indices do not show significant between-seaway structure based on the Bray-Curtis (Fig. 6) or Jacquard dissimilarity (Fig. 7) indices (MANOVA, p > 0.05), as reflected in the structure of the cluster analysis dendrogram (Fig. 8) where most clades comprise sites from more than one seaway.

For most species, subadults were the most abundant class (Table 3), representing approximately two thirds of the total sample. Less than 20% of the 3,398 Epinephelus sp. specimens sampled were classed as adults. The sample mostly comprised juvenile groupers for three species (Epinephelus coeruleopunctatus, Epinephelus fuscoguttatus, and Epinephelus undulosus). Among the rare groupers, the sole representative of Epinephelus heniochus was classed as adult, although the sample size of one does not enable any conclusions to be drawn from this observation. The percentage of adults was less than one third for all the more abundant (n > 50) grouper species, ranging from around 5%–6% (E. coioides, E. maculatus, E. sexfasciatus) to around 26%–30% (E. ongus, E. quoyanus).
The size at first capture (Lc) and size at first maturity (Lm) are important fisheries management parameters. These parameters, along with the maximum size (Lmax) from this study and in FishBase (Froese & Pauly, 2022) are given in Table 4 together the percentage of the sample above Lm and the life stage (juvenile, subadult or adult) at first capture. FishBase, (Froese & Pauly, 2022) had data on length at first maturity (Lm) for five species; however most of the Lm data in Table 4 are from published studies and reports (Ernawati et al., 2021; Fennessy, 2018; Khasanah et al., 2019; Mackie, 2007; Mangi & Roberts, 2006; Mous et al., 2019a; Nair, 2018; Pears et al., 2006; Rhodes et al., 2016; Russell, 2018; Sadovy & To, 2018). No data on Lm were found for four species (E. corallicola, Epinephelus cyanopodus, Epinephelus erythrurus, and Epinephelus melanostigma). Conservation status in Table 5 is from the most recent update of the IUCN Red List (IUCN, 2022), while common names and fisheries uses follow the data in FishBase (Froese & Pauly, 2022).
| No | Species | n | First capture | Lmax | Mean length | First maturity | Reference (Lm) | |||
|---|---|---|---|---|---|---|---|---|---|---|
| TL | Life-stage | This study | FishBase | Lm | % > Lm | |||||
| 1 | Epinephelus areolatus | 688 | 12.44 | Juvenile | 42.00 | 47 | 26.77 | 26.5 | 50.34 | (Mangi & Roberts, 2006) |
| 2 | Epinephelus bleekeri | 72 | 12.21 | Juvenile | 61.21 | 76 | 37.11 | 36 | 50.00 | (Froese & Pauly, 2022) |
| 3 | Epinephelus coeruleopunctatus | 7 | 16.86 | Juvenile | 29.17 | 76 | 23.04 | 31.8 | 0.00 | (Mangi & Roberts, 2006) |
| 4 | Epinephelus coioides | 300 | 13.98 | Juvenile | 98.72 | 120 | 39.32 | 41.3 | 41.55 | (Froese & Pauly, 2022) |
| 5 | Epinephelus corallicola | 55 | 16.13 | No data | 46.45 | 49 | 27.60 | No data | - | |
| 6 | Epinephelus cyanopodus | 2 | 26.50 | No data | 60.48 | 122 | 43.49 | No data | - | |
| 7 | Epinephelus erythrurus | 7 | 16.84 | No data | 31.87 | 45 | 22.61 | No data | - | |
| 8 | Epinephelus fasciatus | 430 | 8.21 | Juvenile | 39.15 | 40 | 22.12 | 18.5 | 82.34 | (Mangi & Roberts, 2006) |
| 9 | Epinephelus faveatus | 229 | 12.45 | Juvenile | 24.46 | 32 | 19.67 | 27 | 0.00 | (Mangi & Roberts, 2006) |
| 10 | Epinephelus fuscoguttatus | 29 | 15.30 | Juvenile | 57.64 | 120 | 30.75 | 40.8 | 9.68 | (Pears et al., 2006) |
| 11 | Epinephelus heniochus | 1 | 25.87 | No data | 25.87 | 35 | 25.87 | 24 | 100.00 | (Mous et al., 2019a) |
| 12 | Epinephelus longispinis | 1 | 27.12 | Juvenile | 27.12 | 55 | 27.12 | 38.5 | 0.00 | (Nair, 2018) |
| 13 | Epinephelus macrospilos | 4 | 22.81 | Juvenile | 34.13 | 51 | 28.47 | < 30 | 50.00 | (Fennessy, 2018) |
| 14 | Epinephelus maculatus | 107 | 11.08 | Juvenile | 45.83 | 60.5 | 29.01 | 30.8 | 38.14 | (Rhodes et al., 2016) |
| 15 | Epinephelus malabaricus | 26 | 22.73 | Juvenile | 65.25 | 234 | 38.73 | 124 | 0.00 | (Mangi & Roberts, 2006) |
| 16 | Epinephelus melanostigma | 3 | 23.86 | No data | 23.98 | 54.9 | 23.93 | No data | - | |
| 17 | Epinephelus miliaris | 9 | 19.78 | Juvenile | 27.76 | 53 | 24.54 | 23 | 50.00 | (Mous et al., 2019b) |
| 18 | Epinephelus ongus | 667 | 13.37 | Juvenile | 42.54 | 40 | 25.25 | 21.1 | 83.23 | (Ernawati et al., 2021) |
| 19 | Epinephelus polyphekadion | 28 | 16.66 | Juvenile | 60.74 | 90 | 33.32 | 36.95 | 33.33 | (Khasanah et al., 2019) |
| 20 | Epinephelus quoyanus | 558 | 6.22 | Juvenile | 36.92 | 40 | 24.15 | 24 | 51.59 | (Sadovy & To, 2018) |
| 21 | Epinephelus rivulatus | 27 | 22.47 | Juvenile | 42.79 | 45 | 30.97 | 19.4 | 100.00 | (Mackie, 2007) |
| 22 | Epinephelus sexfasciatus | 134 | 2.84 | Juvenile | 28.81 | 40 | 16.40 | 15.9 | 51.52 | (Craig, 2018) |
| 23 | Epinephelus undulosus | 6 | 22.05 | Juvenile | 48.82 | 120 | 35.38 | 55 | 0.00 | (Russell, 2018) |
Discussion
The 23 grouper species recorded during this survey comprise nearly 25% of the 93 valid species in the genus Epinephelus according to WoRMS, 44% of the 52 species reported from the Western Pacific (FAO fisheries area 71) and 57.5% of the 40 species reported from Indonesia according to FishBase (Froese & Pauly, 2022). The current IUCN Red List status was LC for 20 of these groupers, with two (the brown-marbled grouper E. fuscoguttatus and the camouflage grouper Epinephelus polyphekadion) listed as vulnerable (VU). While the duskytail grouper (E. bleekeri) is the only species listed as DD, the detailed IUCN assessments also reveal knowledge gaps for several other species, in particular the moderately abundant but little-studied coral grouper E. corallicola.
While all 23 groupers are reported as exploited in FishBase (Table 5), species regarded as major export commodities for the LRFFT from Indonesia include the camouflage grouper E.polyphekadion (Khasanah et al., 2019), the brown-marbled grouper E. fuscoguttatus (Khasanah et al., 2020), and orange-spotted grouper E. coioides (Achmad et al., 2020), two of which (E. fuscoguttatus and E. polyphekadion) have an IUCN Red List conservation status of VU. Collectively, these three species comprised 10.5% of the total sample. For all three species, the length at first capture Lc was less than half the length at first maturity Lm and the mean length was also less than Lm (Table 4). Around 7.8% (Table 3) to 19% (Table 4) of the specimens from these three species were likely mature specimens. In contrast, 78% to 90.5% of E. coioides sampled from deep-water export-oriented grouper and snapper fisheries across Indonesia in 2016 to 2019 were estimated to be mature (Mous et al., 2019b), with a 75% spawning ratio (SPR) for FMA 713 (which comprises the study area), albeit with declining trend in both large mature fishes and SPR, and an increasing trend in the percentage of juveniles caught. Although the number of specimens recorded for these three groupers was comparatively low, possibly because such valuable fish may go to export traders rather than local traders and markets, this contrast indicates that small-scale grouper fisheries selling to local markets could negatively impact the stocks of high-value export commodities, as also reported from Hong Kong (To & Sadovy de Mitcheson, 2009). This exploitation of juvenile high-value groupers can be considered a matter of serious concern for fisheries management, especially as two of these three species are already considered VU under IUCN Red List criteria.
There is an export category of mixed groupers (Khasanah et al., 2020), which may include lesser numbers of the other species identified in this study. Groupers may also be exported fresh, or as processed fisheries products (e.g., frozen or filleted fish) (Nadiarti et al., 2021). However there are extensive anecdotal indications, including many personal observations by the authors, that groupers are widely sold on local markets in Sulawesi as well as in other regions of Indonesia, and are generally smaller-sized than those destined for export markets. This paradigm has been noted in particular in East Kalimantan (Nadiarti et al., 2021), and Central Sulawesi (Ndobe et al., unpublished report to WWF-Indonesia, 2014). As in a study on mostly small-scale fisheries in Eastern Indonesia (Halim et al., 2020), E. areolatus was the most abundant species in this study. Similar to the larger E. coioides, in this study the proportion of adult fish (20%–25%) was much lower than the proportion (97%–99.5%) reported from the above-mentioned study in 2016–2019 by Halim et al. (2020), who also reported an SPR of 49% and stable trends in size. This study appears to confirm the suspicion expressed by Halim et al. (2020) that that catch of juveniles was probably underrepresented in their study due to sampling methods which could miss lower-value fish destined for local consumption. Furthermore, the data indicate a similar situation to that reported for grouper fisheries in Hong Kong, where all sizes have some value and are therefore targeted and retained (To & Sadovy de Mitcheson, 2009).
Of the four species with no data on length at first maturity Lm (Table 4), three (E. cyanopodus, E. erythrurus, and E. melanostigma) can be considered rare or rarely caught; however E. corallicola was found at eight of the 12 sites, including all sites in the Makassar Strait and two each in the Flores Sea and Gulf of Bone, and is listed as a subsistence fishery target in FishBase (Table 5). The life-stage estimation (Table 3) indicates that less than 25% of E. corallicola caught were likely to have spawned. Using the approximation of To & Sadovy de Mitcheson (2009) that Lm50 tends to be close to half of maximum length, this would give Lm ≈ 23–25 cm, indicating that for over half (57%–75%) of the specimens TL might be greater than Lm. This example illustrates the gaps in key life history data for fishes exploited in small-scale and subsistence fisheries where juvenile fish can comprise a considerable proportion of the catch, both in Indonesia (Nadiarti et al., 2021) and in other countries such as Kenya (Mangi & Roberts, 2006).
The greater the gap between size at first capture and size at first maturity, the higher the risk of overfishing (Froese, 2004). In addition to the large proportion of juvenile groupers, observations of mature fish well below the published length at maturity are a cause for concern on at least two fronts. Firstly, early maturity is recognised as a symptom of recruitment overfishing (Achmad et al., 2020; Khasanah et al., 2020; Mous et al., 2019a; To & Sadovy De Mitcheson, 2009). The data from this study indicate a need for management to reduce the proportion of juvenile groupers caught. One potential measure would be to establish minimum legal size regulations for grouper species (or species groups) predominantly exploited for domestic markets and the fisheries that catch them, similar to those proposed for export-oriented grouper fisheries and trade markets (Achmad et al., 2020; Khasanah et al., 2019, 2020). Attention to the gears used is also recommended, as the hook size used in Indonesian hook-and-line grouper fisheries is often very small, thereby increasing the likelihood of catching juvenile fish (Achmad et al., 2020), a fact that many fishermen are aware of (Mous et al., 2019b).
Secondly, there were several observations of mature female groupers that were clearly on the point of spawning, with greatly distended bellies and eggs oozing from the oviduct, including some very small individuals. The condition of these individuals indicates that spawning aggregations are likely being fished, either by chance or by fishers with the local knowledge to identify the time and place of spawning aggregations. This could present a major threat to grouper populations, and hence to the sustainability of the fisheries on which so many people rely for food and livelihoods (Sadovy de Mitcheson et al., 2020). Further research is needed to identify spawning aggregations, together with interventions to protect spawning groupers (e.g., temporal or permanent fishery closures with adequate regulations and enforcement). Raising awareness of the importance of spawning aggregations, and letting juvenile fish grow to maturity before capture is also important, while well-managed protected areas can also play a role (Marcos et al., 2021), even if the location of spawning grounds within them is not yet known.
One major obstacle to sustainable management of groupers is that most effective measures rely on knowledge of species-level biology and ecology. For most of the species in this study there are gaps in this knowledge, partly because of the difficulty of species recognition, which is also one of the reasons for poor data resolution. To support sustainable, ecosystems-based management of grouper stocks, including both export-oriented and domestic-oriented fisheries, we recommend targeted research to fill the gaps in existing knowledge on species-level biology and ecology of Epinephelus groupers, as well as the development and deployment of tools to assist in species recognition, such as the android-based application currently being developed for 18 of the 23 species in this study.